십여 년 전 식물학 분야에서 노벨상을 받는다면 어떤 연구가 받게 될까 하고 동료 교수와 논의한 적이 있다. 당시 개화호르몬을 찾아내면 노벨상을 받을 수 있을 거라 생각했다. 내가 개화 분야의 연구를 하고 1998년 서울대학교에 부임해 왔을 때 내 은사이기도 했던 교수님이 ‘그래, 이제 개화호르몬이 밝혀졌느냐’ 라고 물어볼 정도로 식물학 분야에서 개화호르몬을 찾는 일은 오랫동안 풀지 못한 숙원이었다. 70여 년 동안 풀지 못한 숙제를 푼다면 노벨상을 줘야 하지 않나 라는 게 식물학 분야 연구자들의 일반적인 생각 아니었을까 싶다. 그 개화호르몬, 플로리겐을 밝히는 여정을 쫓아가 보는 것도 나름 의미 있는 일이 아닐까 한다.
개화유도 호르몬, 플로리겐은 1930년대 러시아 과학자 챠일라키얀에 의해 명명되었다. 그는 낮이 짧은 가을에 꽃이 피는 식물-이를 단일식물이라 한다-을 낮이 긴 장일조건에 키우면서 단일 처리를 해준 식물의 잎을 접목해주면 장일조건에서 자라고 있음에도 가지 끝 정단 조직에서 꽃이 피는 현상을 발견하였다(그림). 이는 단일조건에서 자란 잎에서 생성된 개화호르몬이 접목된 가지를 따라서 이동하여 가지 끝 정단조직에 꽃을 피우게 함을 의미한다. 챠일라키얀은 직관적으로 이러한 호르몬의 존재를 상상하고 이를 플로리겐이라 근사하게 이름을 붙인 것이다. 이후 비슷한 접목실험을 통해서 단일식물뿐만 아니라 장일식물에서도 개화를 유도하는 호르몬의 존재가 입증되었고, 심지어 단일식물의 잎을 장일식물에 접목해도 개화가 유도됨을 보여 개화호르몬이 보편성을 가진 분자라는 사실이 확인되었다. 이후 과학자들이 무엇에 관심을 가지게 될지는 불을 보듯 뻔하였다. 전 세계 무수히 많은 과학자가 이 미지의 개화호르몬을 추출하는 작업을 시작했다. 그런데 많은 과학자들이 다양한 생화학적 방법으로 분리를 시도하였지만 플로리겐은 좀처럼 그 모습을 드러내지 않았다. 80년대 초 중반에는 당시의 발전된 물질 추출 기술을 활용하여 마침내 플로리겐을 찾아냈다고 주장하는 논문들이 세계적 명성을 가진 저널들에 심심치 않게 발표되곤 했다. 물론 세월이 흐르면서 그게 아닌 것으로 판정이 나면서 플로리겐을 찾고자 하는 강박증은 점점 더 커지게 되었다.
이런 상황에서 90년대 들어 식물학 분야에서 획기적인 연구 방법이 개발되게 된다. 모든 연구자들이 함께 실험재료로 사용하는 모델 식물, 애기장대의 발굴과 분자유전학적인 도구들이 개발되기 시작한 것이다. 생물학 연구에서 모델 생물의 개발은 대단히 중요하다. 같은 생물 종을 실험재료로 사용하게 되면 여러 과학자들이 그 생물을 연구하기 위해 개발한 각종 방법들이 집적되게 되고 결과적으로 생명현상을 이해하는 연구의 속도가 대단히 빨라지게 되는 것이다. 그런 모델생물로 가장 모범적인 사례가 된 것이 대장균과 효모이고, 동물로는 초파리, 예쁜꼬마선충, 쥐 등이 모델생물로 구축되었다. 식물학 분야에서도 모델 식물을 찾는 노력이 있어서, 80년대에는 페튜니아를 모델 식물로 사용하자고 제안되기도 했다. 그러나 작은 크기, 빠른 세대 기간, 아주 작은 유전체의 크기 등의 여러 가지 장점 때문에 애기장대가 90년대 이후 빠른 속도로 식물학 분야의 ‘워너비’가 되었다. 애기장대 모델 식물의 구축으로 개화유도 분야를 연구하던 과학자들은 70년간의 미스테리를 풀 수 있는 방법을 갖게 되었다고 좋아했다. 그동안 플로리겐을 찾기 위해 활용했던 생화학적인 방법 대신에 유전학적인 방법을 활용할 수 있게 된 것이다. 원리는 이렇다. 개화되지 않는 돌연변이체를 선별하고, 이 돌연변이체에서 망가진 유전자가 무엇인지 확인하면, 그 유전자의 생화학적인 기능을 추론해서 플로리겐을 찾게 된다는 것이다. 이 간단한 아이디어를 쉽게 적용할 수 있는 식물이 모델 식물인 애기장대인 것이다.
이러한 이론적 근거를 바탕으로 많은 과학자들은 애기장대에서 개화 돌연변이체를 선별하게 된다. 이상적으로는 개화가 아예 안 되는 돌연변이체를 찾으면 플로리겐의 규명에 더 가까이 다가가는 일이겠지만 당시 유전학자들의 손에 그러한 돌연변이체는 얻어지지 않았다. 대신 개화가 현저히 지연되는 돌연변이체들이 십여 개 이상 동정 되었다. 이를 개화지연 돌연변이체라 하는데 각각의 돌연변이체 이름을 멋없는 fa, fb, fc, fd, (flowering) 등등 일련 번호 붙이듯이 명명하였다. 후에 개화유전자들이 하나씩 규명되면서 낭만이라고는 찾아볼 수 없는 이름을 아쉬워했던 과학자들이 이들의 이름을 다시 FLOWERING LOCUS D(FD), FLOWERING LOCUS T(FT) 등으로 개명하게 된다. 개인적으로는 유전학의 전통을 따르지 않는 유전자 명명 방식이 마음에 들지 않아 나는 곧 죽어도 FT, FD라고 부르고 있다. 최모 여사처럼 마음에 안 든다고 제멋대로 이름을 고치면 안된다는 규칙이 유전학의 전통이기 때문이다.
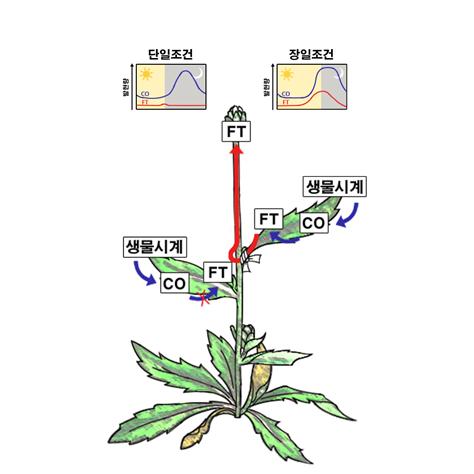
어쨌든 개화지연 돌연변이체들로부터 개화유전자들이 하나씩 동정되기 시작한다. 최초로 동정된 개화유전자는 루미니디펜던스(LUMINIDEPENDENS)라는 꽤 그럴듯한 이름이 붙은 유전자인데 필자에 의해 1993년에 논문으로 발표되었다. 이후 개화유전자들이 계속 동정 되었지만 플로리겐이 무엇인지 짐작조차 할 수 없는 상태가 계속되었다. 2002년에는 애기장대가 장일조건과 단일조건을 어떻게 다르게 인지하여 장일조건에서는 꽃이 빨리 피고 단일조건에서는 꽃이 늦게 피는지 분자 수준에서 이해하게 됐다. 간략히 설명하면, 개화지연 돌연변이체로 동정된 콘스탄스(constans)와 ft는 장일조건에서는 개화가 야생종에 비해 늦게 일어나지만, 단일조건에서는 야생종과 같은 시기에 꽃이 피는 표현형을 보인다. 말하자면 광주기 신호를 인지하지 못하는 돌연변이체들이다. 그런데 이 유전자들을 동정해서 분석해 보니 콘스탄스나 FT 유전자의 발현량을 인위적으로 증가시키면 개화가 대단히 빨라진다는 사실을 알게 됐다. 더구나 콘스탄스는 유전자 발현이 생물학적 시계에 의해 조절되는데 환경이 단일조건이건 장일조건이건 상관없이 새벽 동틀 때 일정량의 발현을 보이고 이후 낮 동안에 발현이 현격히 줄어들었다가 늦은 오후에 증가하기 시작하여 14시간 이후가 되면 높은 발현량을 보였다. 비록 생물학적 시계에 의해 콘스탄스의 발현이 비슷하게 조절되지만 장일조건과 단일조건에서는 분자·생리학적 차이가 발생하게 된다. 즉 장일조건에서는 여전히 태양이 떠 있는 낮에 콘스탄스의 발현량이 높고, 단일조건에서는 해가 지고 밤이 되었을 때 콘스탄스의 발현량이 높다는 생리적 차이가 발생하게 된다(그림). 이러한 결과는 과거 생리학적인 실험을 통해 알고 있었던 빛과 생물학적 시계가 동시에 작용할 때 꽃이 핀다는 시간적 일치성 이론과 잘 부합하는 현상인 셈이다. 실제로 콘스탄스는 FT 유전자 발현을 촉진하는 전사조절 단백질이고 빛이 있을 때 역치값 이상의 콘스탄스 단백질이 존재하면, 즉 장일조건이면, FT의 발현양이 많아져서 개화를 촉진하게 된다. 반면 단일조건에서는 빛이 없는 밤에 콘스탄스 발현이 증가하게 되는데 이때는 콘스탄스 단백질이 생성되는 족족 분해되어 없어지기 때문에 FT 발현을 매개하지 못하게 된다(그림). 이러한 간단한 실험 결과를 통해 그간에 알고 있었던 개화유전자들이 어떻게 생물학적인 시계에 의해 조절되는지, 나아가서 식물이 어떻게 장일조건과 단일조건을 구분하여 개화 시기를 촉진하거나 늦추는지를 명확하게 이해하게 됐다. 심지어 2003년에는 단일식물인 벼에서도 콘스탄스-FT 유전자 모듈에 의해 개화 시기가 결정되는데, 장일조건의 벼 콘스탄스는 애기장대에서와 달리 벼 FT 발현을 억제하는 억제인자로 작용하기 때문에 장일조건에서 개화가 지연된다는 사실도 밝혀지게 된다.
이처럼 광주기의 분자 메커니즘이 밝혀졌고, 콘스탄스-FT 유전자 모듈이 거의 모든 식물에 보존되어 있다는 사실이 밝혀졌음에도 여전히 플로리겐의 정체는 밝혀지지 않고 있었다. 접목에 의해서 이동되면서 모든 식물에 보편적으로 존재하는 개화호르몬, 그러한 특성에 맞는 호르몬이 무엇인지 오리무중인 상황에서 FT가 호르몬으로 작용하지 않을까 짐작하는 과학자들이 등장하게 된다. 그동안 생화학적인 실험을 하면서 연구자들이 플로리겐의 후보로 찾은 분자는 우리 인간에게서 발견되는 여러 가지 호르몬처럼 크기가 작고 이동이 쉬운 화학 분자를 줄곧 생각해 왔다. 그런데 FT는 유전자 산물로 단백질이다. 단백질은 일반적인 호르몬과 비교했을 때 그 크기가 어마어마하게 큰 분자이다. 이렇게 큰 분자가 접목실험을 통해서 한 가지에서 다른 가지로 이동한다는 것은 쉽게 상상할 수 있는 일이 아니었다. 따라서 과거 70년간의 생화학적 실험에서 플로리겐의 추출 및 분리를 시도할 때 단백질이나 핵산은 아예 제거해 버리는 분자 덩어리 속에 들어가 있었던 것이다. 따라서 플로리겐이 단백질과 같이 큰 분자라면 그동안 왜 플로리겐을 추출하지 못했던가를 잘 설명할 수 있게 된다. 이러한 가설은 실제 확인을 해보니 사실에 가까웠다. 그래서 마침내 2007년 FT 단백질이 70년간의 미스테리였던 개화호르몬, 플로리겐이라 주장하는 논문이 발표되었다. 이후 9년의 세월이 지났는데 FT=플로리겐이라는 가설이 깨어지지 않고 유효한 것으로 보아 FT 단백질이 우리가 찾던 플로리겐이 맞는 듯하다. 그런데 플로리겐을 찾으면 노벨상을 탈 수 있을 것 같았던 기대는 허물어지고 말았다. FT 유전자는 이미 15년도 더 전에 밝혀져 있던 유전자이고, 그게 플로리겐이라고 밝혀지는 과정이 전혀 드라마틱하지 않아 김샌 콜라를 손에 쥔 기분이었기 때문이다. 참으로 안타까운 발견이 아닐 수 없다.
저작권자 © 포항공대신문 무단전재 및 재배포 금지
